



スキーマ理論
動きに関した運動プログラムの作成とその記憶ということを考えた場合、新しい動きの一つ一つに対して、その都度運動プログラムを作成し、それを記憶していくとなると、その数は無数になってしまうと考えられ、現実的ではない。そこで運動行動に関した運動プログラムの理解に、スキーマの概念が取り入れられた。この考え方によって、運動行動においての汎用性のある運動プログラムに関する枠組みがつくられる。つまり運動行動に関し、全体の基本的な枠組みは同じで、個々の内容を変えることによって、さまざまな状況に対応した運動プログラムがつくられ、これにかかる労力を減らすことができる。これが運動プログラムにおけるスキーマ理論である。
スキーマ(図式という意味)とは、特定の概念を表象するための構造化された知識の集合と考えられている。認知機能におけるスキーマ理論とは、外界の事物を理解する場合、たとえばイスであれば、キャスターがついて動ける一人用のものもあれば、何人かが座れる長イスもあり、また座面が柔らかくクッション性に富んだものもあれば、比較的かたいものもあり、デザインもいろいろで、さらに会議室にあるものは一人掛けのもの、待合室であれば長イスなどいろいろであり、それらが「イス」という一つにカテゴリーとして認識されていて、これが知識構造となって「イス」というものが理解されるというような考え方をいう。ちなみにこれらの知識は、意味記憶またはエピソード記憶として長期記憶化されたものである。
次に一般的に運動行動がなされる場合に、入手される情報として、A初期条件(運動行動を開始する直前の環境などについての情報)、B反応明細(一般運動プログラムをどのように実行するかを決定する力やスピードなどのパラメーターの情報)、C感覚経過(運動行動に伴って生起する感覚情報)、D反応結果(運動行動遂行の結果ならびにその結果とフィードバック情報との誤差)がある。運動技能学習にスキーマ概念を持ち込んだリチャード・A・シュミットの理論では、「再生スキーマ」と「再認スキーマ」とから構成される「運動反応スキーマ」を想定している。スキーマ理論における運動制御の流れは、まず目標となる運動行動の結果と初期条件が入力されると、再生スキーマによって反応明細が生み出され(これは「結果」から必要な「動き」が導かれる形なので、逆モデルの構造となっている)、それにもとづいて運動プログラムが筋肉に指令を発し運動行動が実行される。それと同時に再認スキーマは、誤差検出のための基準として働く期待される感覚経過として、目標としている運動行動を実行したらこのような動きの感覚がフィードバックとして得られるであろうという筋感覚的(体性感覚的)な予期、イメージと、視覚的あるいは聴覚的予期、イメージとを生み出し、運動行動の実行によって生じた感覚経過が、一つは自己受容感覚(体性感覚)として前者と比較され、一つは外部受容感覚(視覚と聴覚ということ)として後者と比較され、もう一つは最終的に得られた運動行動の結果として、はじめに入力された目標と比較され運動反応スキーマをより正確なものとするために使われる(目標とする結果と実際に得らえた結果との誤差を最小限にしていく)。これらの過程を繰り返し行うことで、新しい運動反応スキーマが形成されるということになる(1)(図6)。
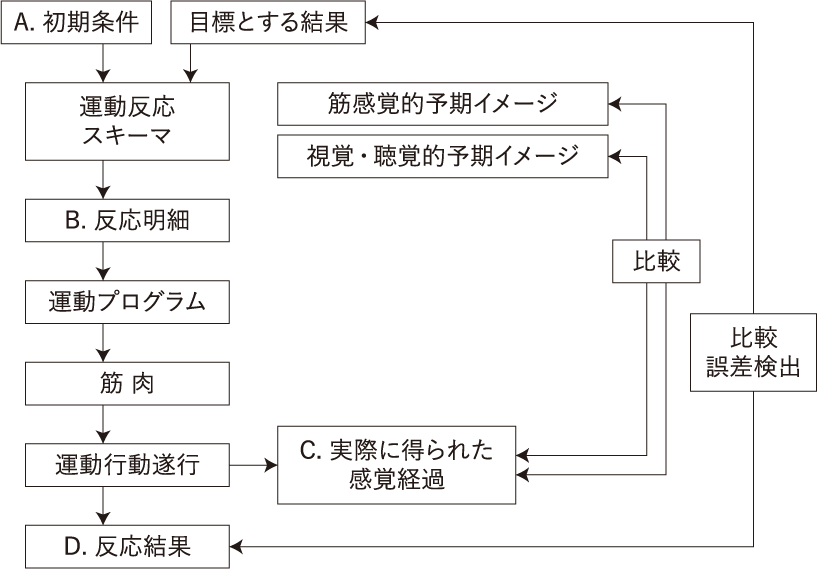
このABCDを、さらにこまかく見ていくと、次のような情報になる。a.行動を起こす前の状況、つまりその運動行動を行う目的や意図が生じた状況、環境ということ。b.運動行動を行うことによって予測され期待される結果。c.行おうとする運動行動の運動指令、反応明細といわれる、つまりどのように動かすかということ。この過程によって、用意されている運動プログラムが筋肉に指令を発し、運動行動が実行される。d.運動の実行そのもの。e.運動行動が実行される際の直前の感覚、体性感覚、視覚、聴覚などによる、これは無意識に知覚される場合もある。f.その運動行動の実行に伴って生じた手足の体性感覚、視覚、聴覚による感覚。これらは運動行動に伴って生じる感覚として常に予測されていて、実際に生じた感覚と比較照合される。もし予期された感覚と異なっていれば、その時点で運動行動の継続がうまくいかなくなるか、中止される(動きが止まる)。またこれらの感覚の知覚によって、実際に体がどう動いているか(実際の運動軌道)を知覚、認識できる。g.運動行動によって実際に得られた結果。h.運動行動の目的、意図(目標とした結果)と実際に得られた結果との誤差の評価、となる。
運動行動に伴う感覚とは、たとえばATMなどで暗証番号を押す際に、平面のタッチパネル式でなくボタン式の場合だと、ボタンを押した時の指の感覚というのは、記憶にもとづいて無意識に知覚されているが、ATMの機種の違いによってはボタンを押した時の感覚が、記憶にもとづいて予測されていた感覚と微妙に違うものもあり、いつもと違ったボタン押しの感覚が得られると、押した時に違和感が生じ、それ以上ボタンを押すことをためらい、やめてしまうということもあるだろう(押したときの感覚が以前押した時のものと同じなら意識されない、もし違った感覚であった場合、違和感として意識される)。
バレーボールでサーブを打つ時、ボールをこう持ってこう打とうという運動指令を発し、今までと同じような体の動きで、同じように力を加えてボールを手で打ち、今までと同じような手の感覚が得られたとしても、ボールが同じ軌道を描いて飛んでいくとは限らない。会場の温度、湿度、空調の風向きなどで、ボールの軌道は(微妙に)変化する。体の動きとボールに与える力が今までと同じ、つまり運動指令と得られた感覚が同じであったとしても、得られた結果は変わる可能性があるということで、この場合、期待された結果と実際の結果との差異、つまり誤差が生じることになる。
体操選手が競技に入る前に、手指の感覚や、鉄棒などの器具と手の接触の際の感覚を何回も確認するという話を聞いたことがある。会場の湿度や、滑り止めの粉の量やつけ方などによって、得られる手の感覚が違ってくる可能性がある。実際の競技に入る直前の、得られるはずの手の感覚が少しでも練習のときと違うと、今までと同じ動きができなくなる可能性がある。体操選手は今までの練習の時と同じ感覚を得るために、滑り止めの量を調節し、何度も手の感覚を確認しているのだろう。
動きにおけるフィードバック制御
動きにおけるフィードバック制御とは、まず動きが実行されてから、その動きに伴う感覚を知覚し、目標とする軌道とずれていれば、あとからその動きを修正しようとする機能のことだが、感覚の知覚には、はやくても50~150ミリ秒かかるため、軌道のずれの修正はそのぶん遅くなる。かりに50ミリ秒で知覚したとして、それを行動に反映させるためには、反応時間の測定(光などの視覚刺激が見えたら素早くボタンを押すなど)の時のように、最も速い反応性の動きであっても、100ミリ秒以内では反応できず、反応時間はだいたい150~250ミリ秒といわれるので、フィードバック制御においても、動きが修正されるまでには同様の時間はかかると思われ、目標とする正確な軌道への修正に利用するには、やや遅くなってしまう。それでは動きにおけるフィードフォワード制御というのはどうなっているのかを次に述べる。
小脳と内部モデル、フィードフォワード制御
小脳は、日常の文脈的な動きなどに恒常的に関与しているといわれる。小脳は運動に関与することがよく知られていて、小脳機能としては、動きをスムーズにしたり、目標物に手をまっすぐに正確に持っていったりと、メリハリのある行動をつくりだすことに関係している。他の機能として、距離や時間に対する感覚の認識がある。目標物に手を伸ばしてそれをピタッとつかむことができるのは、小脳が手と目標物との距離を認識し機能するからと考えられる。小脳は動きに関与しているが、大脳皮質の運動関連領域と直接関連している領域は小脳全体の20~30パーセント程度といわれ、運動野以外で前頭前野、頭頂連合野、大脳基底核などと関連がある。小脳は動きだけではなく、実は感覚にも密接に関係している。もともと感覚と動きというのは密接に関係していて、一般的にまず何らかの感覚があって、それに対する動きがあって、さらにその動きに伴う感覚があるという一つのパターンがある。動きと感覚というのは、常に同時的なものともいえる。動く時にはいつもそれに伴って得られる感覚を予期、予測している。ちなみに動きの認識というのは、感覚の知覚のことである。運動感覚というのは、動きに伴って起きる感覚、つまり筋紡錘や腱や関節からの深部感覚と視覚や聴覚でつくられるもので、基本的に感覚なのである。つまり動きの認識は「動いているという感覚」の認識のことである。
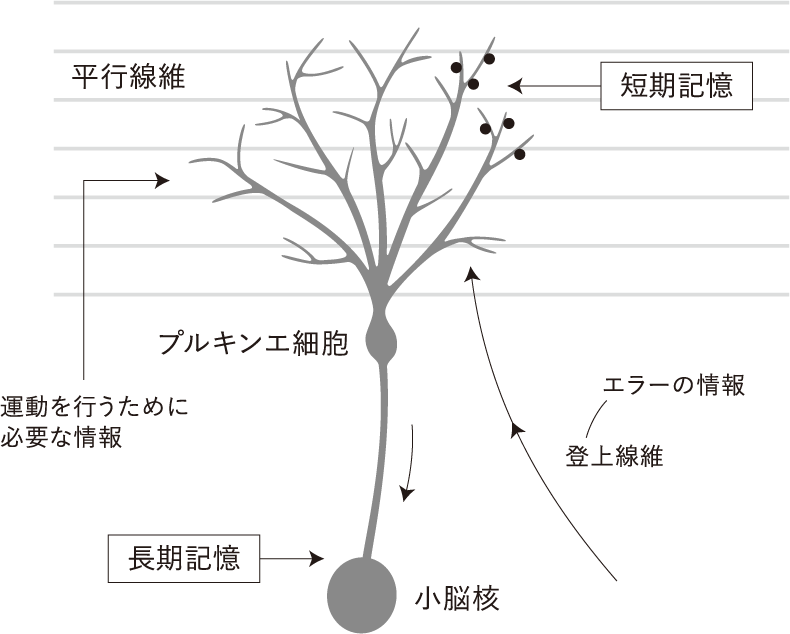
動きに重要な小脳のプルキンエ細胞への神経連絡には、顆粒細胞の軸索による平行線維からの入力と、登上線維からの入力がある。1個のプルキンエ細胞に対して、およそ20万本の平行線維がシナプスを形成しているが、登上線維からの入力は1本である。この多数の平行線維は、動きの直前(約20ミリ秒前)の情報に関係しているといわれる。次にプルキンエ細胞からの出力によって行動が起こされるわけだが、そのあとこの動きが状況にそぐわないものであれば、エラーの信号が登上線維からプルキンエ細胞に伝わり、平行線維とプルキンエ細胞との間に長期抑圧という機構がはたらき、平行線維とプルキンエ細胞との間に動きに関する短期記憶ができる。この際に短期記憶は数時間で消えるが、何回も、何日も動きを繰り返すことによって、必要な動きが長期記憶化される。このとき動きの記憶は小脳核というところに移動し、長期増強によって長期記憶となるという(図7)。この長期記憶化した動きの記憶が、一般に手続き記憶といわれる動きの記憶のことである。
手続き記憶は動きに関してだけではなく知覚や認知的なことに関してもいわれる。手続き記憶は言語化しにくい非宣言的な記憶といわれる。たしかに動きというのは、言語化はしにくいかもしれない。ただもともと言語というものは、すべてを表現するには完璧とはいえないとも考えられている。身近な例では、料理などの味の表現というのは、甘いとかからいとか、さっぱりしておいしいとか、いくつかの表現に限られている。コクがあるとか、深みがあるとかいわれるが、それではコクとか深みとはさらにどのような感覚をいうのかということになると、これ以上の表現は難しい。桃も夏みかんも、味はどちらも甘酸っぱいと表現されるが、両者は全く違う味であり、甘みが強いとか、酸味が強いとか、さっぱりしているなどで表現されるかもしれないが、厳密に質的な味の違いの表現というのは言葉では難しい。結局は前に経験した味の記憶などから、自分なりに理解するしかないだろう。
ちなみに手続き記憶としてよく自動車の運転がいわれる。自動車のレバーをドライブにいれるとか、アクセルを踏むなどで、これらは車を動かすための一種の道具であり、意味記憶によって、レバーとかアクセルを操作するということになる。このこと自体は道具使用ということと同様だが、運転することにおいては、これらの使用における相互作用と全体的な統合が必要であり、これが運転の技術すなわち手
続き記憶ということになるわけである。
小脳には内部モデルがあるといわれているが、そのことをあらわす次のような実験結果がある(11)。小脳に関しての実験で、周期的に現れる視覚刺激に同期して眼を動かすようにサルを訓練することに成功したというこの実験結果によると、小脳歯状核神経には、a.特定方向の運動の前に活動するもの、b.方向に関係なく運動の前に活動するもの、c.運動の直後に活動するものがあることがわかったとのことである。aのタイプは次に行う運動のタイミングとよく相関した活動を示し、運動の制御に直接かかわっているものと考えられ、bのタイプは運動そのものよりも周期的に現れる標的のタイミングに一致した活動を示し、標的自体の内部モデルを表象しているものと考えられ、cのタイプは標的と運動の時間ずれ(エラー)とよく相関した活動を示し、同期運動の時間誤差を検出することに関与すると考えられたとのことだった。この場合のbの標的のタイミングに一致した活動の内部モデルというのは、外部世界を脳内に表象するという意味での内部モデルということとなる。ここでaの運動のタイミングと相関していて、運動の制御に直接かかわっている機能というのも、運動制御における内部モデルといえるのではないかと考えられる。aは確かに脳内部の表象ではあるが、もとになっているのは感覚の知覚の記憶である。このことからいえるのは、小脳にはさまざまな感覚の記憶が貯蔵されていて、現在生じている感覚の次に起きるはずの感覚もすでに経験によって記憶されているので、その起きるはずの感覚に合わせた動きを、前もってつくる、準備することができるということとなり、これがフィードフォワードという動きの制御ということになる。小脳は経験によって得られたさまざまな感覚に伴う知覚を組織化していて、それらの感覚の記憶から、次の感覚を予測し、それに見合った動きをつくりだすというフィードフォワードの機能によって、リアルタイムに、次の感覚に一致した(生じるはずの感覚に遅れない)素早い運動生成が可能となっている。このように感覚の知覚から次の動きを生成していく過程において、行動の経験によって学習された動きの記憶は内部モデルとして小脳にあり、予測された感覚に対する動きの生成において、この内部モデルがもとになって動きが発現されていると考えられる。これが小脳における動きの発現に関するフィードフォワード機構ということとなる。
リズムに合わせて踊ったりする時のように、リズムを感じているとき、脳内では周期的に繰り返されるパターンを再現した内部モデルがつくられていて、これは小脳において外界から入力される周期的なリズムの内部モデルが生成されているということになり、これによって、そのリズムに合わせて踊るなどの行動ができるということのようである。これも小脳における動きのフィードフォワード機能によると考えられる。
またマウスを動かしてカーソルで軌道を作成する実験において、マウスを直線的に動かすと、直線ではなくカーソルの軌道がずれていくという条件で、それぞれ異なった軌道をつくる2種類のマウスを動かすことによって、カーソルの動きを学習させてみる。動きの軌道の学習を行っている間は、小脳の短期記憶で活動する領域が活性化し、学習が進むにつれ、活性化の領域が小脳の歯状核(小脳核の一種)に限られていくようになるという。そして二つのマウスの動かし方の記憶は、それぞれ別々の歯状核の領域で活性化するという。これはマウスの動かし方の記憶がそれぞれ長期記憶となって歯状核に移動するということを意味し、このことはそれぞれのマウスの動かし方の記憶が長期記憶として歯状核のそれぞれの部位に記憶され、保存されることを意味することになるという。そしてこの動きの長期記憶というのが、動きの内部モデルということになるわけである。つまり二つの異なったカーソルの動き方をするマウスをそれぞれ動かす際の、動かし方の内部モデルというのが、小脳の歯状核の中の、それぞれの違った部位に保存されるということになるという。
さらにこれらのことから、小脳のフィードフォワード機能は、意味的な解釈としては、行動における逆モデルを解いているということになると考えられる。先ほどの実験での、視覚刺激に同期した眼の移動においても、マウスの動かし方においても、内部モデルの順逆モデルという点では、逆モデルを解くということになる。つまり眼の動きを視覚刺激に同期させるには、どのように眼を動かしたらいいのか、動かすタイミングと眼の方向を逆モデルで解いてタイミングを合わせるという意味合いとなり、また直線的に動かすと直線軌道を描かないマウスで直線を描くには、どのようにマウスを動かしたらいいのかということを、目標軌道からマウスの動かし方の軌道を逆モデルとして解いて、マウスを動かすということになるからである。したがって小脳に存在するといわれる内部モデルは、行動における逆モデルを解くという点で機能していると考えられる。
いままでの内容は実験結果によるものだが、日常生活においても小脳が機能していることが理解できる場面がたくさんある。たとえば、瓶を上に投げ上げて、落ちてくるところをうまくキャッチするためには、小脳がはたらいている。上に投げ上げられた瓶は重力にひっぱられてある時点から落ちてくるが、その落ちてくる軌道を小脳が予測できれば、タイミングよく手でキャッチすることができる。これも落ちてくる瓶をうまくキャッチするにはどうしたらいいかというその瞬間の認識において、逆モデルを解いているということになる。ものを空中に投げ上げてから落ちてくるときの物体の軌道と、それにかかる時間というのは、おそらくは小さいころからの知覚の経験と学習によって、小脳に記憶されていて、さらにおちてくる物体をキャッチする行為も、やはり同様に多くの経験による学習によって、小脳に長期記憶として記憶されていると考えられる。つまりこれらは小脳における内部モデルとしての記憶ということになっていて、小脳は瓶が落ちてくる状況から、いつ手をだしてキャッチしたらいいのかという行動のタイミングをはかっている、予測しているのではないだろうか。このタイミングは学習によって小脳に記憶されていると考えられる。またその瓶を手でつかんで把持するという、手の動きに関して、その全過程を、どれくらいの時間内に完結させるかという、瞬間的な動きであれば数百ミリ秒程度と思われるが、動き全体に必要な最小限の時間間隔というのも小脳が把握していて決めているのではないだろうか。その時の状況によって、思ったよりも素早く動作を完結させなければならない状況もあると思われ、その場合には素早い動作をつくりだすように機能し、小脳はその動きの時間幅の調節にも関与しているように思われる。またもちろん、小脳は手からその物体までの距離を認識し、その物体に手を伸ばして正確に手でつかむ行為にも関与している。このように、小脳は動きを前もって予測し、フィードフォワード機能によって、その状況において遅れのない動きをつくりだすことに寄与しているのだろう。
参考文献
- 1)『人間の運動学 ―ヒューマン・キネシオロジー』
宮本省三 (著)、 八坂一彦 (著)、 平谷尚大 (著)、 田渕充勇 (著)、 園田義顕 (著)
協同医書出版社 2016年 - 11)「霊長類の小脳深部核における運動同期を制御する神経信号」
岡田研一、竹谷竜二、田中正樹
ネイチャーコミュニケーションズ13/2022年
| 動きと意識 【全7回】 | 公開日 |
|---|---|
| (その1)Ⅰ 動きと意識のダイナミクス | 2025年6月30日 |
| (その2)Ⅰ 動きと意識のダイナミクス | 2025年7月30日 |
| (その3)Ⅰ 動きと意識のダイナミクス | 2025年8月29日 |
| (その4)Ⅰ 動きと意識のダイナミクス | 2025年9月30日 |
| (その5)Ⅱ 脳と運動の相互作用 | 2025年10月31日 |
| (その6)Ⅱ 脳と運動の相互作用 | 2025年11月30日 |
| (その7)Ⅱ 脳と運動の相互作用 | 2025年12月26日 |